2. Reagents
All reagents and solvents used in this study were of analytical grade, sourced from British Drug House, Poole, England.
2.1. Experimental Design
This study aimed to assess the biochemical and histopathological effects of a mixture of dichlorvos, dimethoate, and cypermethrin on female rats across various biological stages: exposure, mating, pregnancy, and lactation. Analytical-grade dichlorvos, dimethoate, and cypermethrin were obtained from reputable suppliers to ensure consistency and reliability in the experimental processes.
A total of 64 female rats and 16 male rats, matched for strain, weight, and age, were acquired for the experiment. The animals underwent a one-week acclimatization period to reduce stress-induced variability and ensure optimal health prior to the study. The female rats were randomly assigned into eight groups (A-H), each comprising eight rats. Each group was exposed to predetermined doses of the chemical mixture, administered via a controlled route to ensure uniform exposure. The exposure phase spanned 28 days, during which the health and behavior of the rats were closely monitored. After this phase, four rats per group (n=4) were randomly selected and sacrificed for biochemical and histopathological assessments, providing baseline data on the effects of the chemical mixture during exposure.
Following the initial sacrifice, the remaining four rats in each group were paired with two male rats per group to facilitate mating. Behavioral monitoring confirmed successful copulation, after which the males were removed to minimize stress and promote the females’ well-being. The pregnant females were observed throughout gestation for any physiological or behavioral alterations. Post-delivery, the lactating females were allowed to nurse their pups for 10 days. During this period, maternal care behaviors were evaluated, and pup health and growth were recorded to identify potential transgenerational impacts of the chemical exposure.
At the end of the lactation period, the lactating females were sacrificed, and detailed biochemical and histopathological analyses were conducted. Data from both the exposure and lactation phases were subjected to rigorous statistical analysis to identify dose-dependent effects, significant trends, and potential long-term toxicological impacts. These findings contribute to a comprehensive understanding of the biochemical and physiological effects of dichlorvos, dimethoate, and cypermethrin on female rats across critical biological stages, highlighting their toxicological implications.
2.2. Animals
Adult virgin female albino Wistar rats, aged 2-3 months and weighing 190-200 g, were selected for this study. The animals were sourced from the Animal House of the Federal University of Petroleum Resources, Effurun. Upon arrival, they were housed in well-ventilated cages, with five rats per cage, under hygienic and controlled environmental conditions. The room temperature was maintained at 25 ± 2°C, with relative humidity at 50 ± 10% and a photoperiod of 12 hours of light and 12 hours of darkness.
Throughout the study, the rats were fed a standard pelleted rodent diet and provided with clean drinking water ad libitum. Bedding materials were routinely replaced, and cages were cleaned regularly to uphold high hygienic standards, ensuring the animals remained healthy and stress-free. These measures minimized external variables and supported the generation of consistent and reliable experimental outcomes. All animal handling and experimental procedures were conducted in strict compliance with ethical guidelines for the care and use of laboratory animals.
2.3. Dose Levels
The study involved the use of three pesticides: dichlorvos, dimethoate, and cypermethrin, all sourced from Hubei Sanonda Co. Ltd, China. These agrochemicals were purchased from a licensed agrochemical distributor and diluted with clean water to reflect concentrations typically used in domestic applications. The dose combinations and pesticide exposures for each experimental group were administered based on the manufacturer's instructions, as follows:
Group A (Control): Exposed to water sprayed under identical conditions without any pesticide.
Group B: Exposed to dichlorvos, diluted at a 1:1 ratio.
Group C:Exposed to dimethoate, diluted at a 1:1 ratio.
Group D:Exposed to cypermethrin, diluted at a 1:1 ratio.
Group E: Exposed to a mixture of dichlorvos and dimethoate, prepared in a 1:0.5:0.5 ratio.
Group F:Exposed to a mixture of dichlorvos and cypermethrin, prepared in a 1:0.5:0.5 ratio.
Group G:Exposed to a mixture of dimethoate and cypermethrin, prepared in a 1:0.5:0.5 ratio.
Group H:Exposed to a mixture of all three pesticides (dichlorvos, dimethoate, and cypermethrin), prepared in a 1:0.33:0.33:0.33 ratio.
The solutions were freshly prepared daily and sprayed into a poorly ventilated compartment containing the animal cages. This setup simulated domestic pesticide application in confined spaces. The exposure lasted for 28 consecutive days, during which the health and behavior of the animals were carefully monitored.
This exposure method ensured that the study reflected real-life conditions, enabling an accurate assessment of the toxicological effects of dichlorvos, dimethoate, and cypermethrin individually and in combination. The control group (Group A) provided a baseline for comparison, as these animals were exposed only to water under the same experimental conditions.
2.4. Mating and Fertility Assessment
On post-treatment day (PTD) 29, the remaining treated female rats (n = 4 per group) were cohabited with proven fertile adult male rats at a 2:1 female-to-male ratio to assess mating and fertility outcomes. Vaginal smears were collected daily from the cohabited females, and the presence of sperm in the vaginal smears was used as confirmation of successful copulation. The day sperm-positive smears were detected was designated as gestational day zero (GD 0).
The duration required for each female to produce sperm-positive smears was recorded to evaluate potential effects of the pesticide treatments on mating efficiency and fertility. This approach enabled the identification of any treatment-related delays or impairments in the reproductive processes of the treated female rats.
2.5. Blood Sampling
At the conclusion of the 28-day treatment period, blood samples were collected from the treated female rats to facilitate hematological and hormonal assessments. Blood sampling was performed under light chloroform anesthesia, with samples obtained from the dorsal aorta of four females per group.
The collected blood was divided into heparinized and non-heparinized tubes. Blood in the heparinized tubes was used for hematological analyses to examine the impact of the treatments on blood parameters. Non-heparinized blood samples were centrifuged at 4000 rpm for 15 minutes to separate serum. The extracted serum was stored and subsequently used for hormonal assays to assess reproductive and physiological health.
2.6. Hormonal Assay
The determination of serum hormone levels was carried out using standard methodologies as follows:
2.6.1. Follicle-Stimulating Hormone (FSH)
FSH concentration in serum was determined using the sandwich enzyme-linked immunosorbent assay (ELISA) method. The FSH in the sample bound to specific antibodies coated on the microplate, followed by the addition of an enzyme-labeled secondary antibody. A substrate solution was then added, and the intensity of the color developed was directly proportional to the FSH concentration in the serum
| [12] | Burtis, C. A., Ashwood, E. R., & Bruns, D. E. (2012). Tietz textbook of clinical chemistry and molecular diagnostics. Elsevier. |
[12]
.
2.6.2. Luteinizing Hormone (LH)
The serum concentration of LH was measured using the chemiluminescent immunoassay (CLIA) method. Labeled antibodies reacted with LH in the sample, and the resulting chemiluminescence signal was measured. The intensity of the light emitted was proportional to the LH concentration
| [13] | Diamandis, E. P., & Christopoulos, T. K. (1996). Immunoassay. Academic Press. |
[13]
.
2.6.3. Prolactin
Prolactin levels were quantified using the electrochemiluminescence immunoassay (ECLIA) technique. In this method, prolactin in the serum bound to specific antibodies labeled with a chemiluminescent tag. When stimulated by an electric current, the tagged antibody emitted light, which was measured to determine prolactin levels
| [14] | Klee, G. G. (2000). Estrogen and related compounds. Clinical Chemistry. |
[14]
.
2.6.4. Estradiol (E2)
Estradiol determination was conducted using the radioimmunoassay (RIA) or ELISA method. In the ELISA approach, estradiol in the serum competed with an enzyme-labeled estradiol analog for binding to specific antibodies. The resultant color intensity after substrate addition was inversely proportional to the estradiol concentration
.
2.6.5. Progesterone
Progesterone levels in serum were assessed using the competitive ELISA method. Serum progesterone competed with enzyme-labeled progesterone for binding to specific antibodies immobilized on the assay plate. The intensity of the color developed was inversely proportional to the progesterone concentration
.
2.6.6. Testosterone
Testosterone levels were determined using the enzyme-linked immunosorbent assay (ELISA) method. In this procedure, testosterone in the serum competed with enzyme-labeled testosterone for a limited number of antibody-binding sites on the assay plate. The color intensity developed after the substrate reaction was inversely proportional to the testosterone concentration in the sample
| [4] | George, J., Shukla, Y., & Tandon, S. K. (2011). Cypermethrin-induced ovarian dysfunction and oxidative stress: A review. Journal of Reproductive Toxicology, 5(2), 102-109. |
[4]
.
2.7. Statistical Analyses
Data were analyzed using one-way analysis of variance (ANOVA) to assess the effects of pesticide exposure on hormonal parameters. Post-hoc comparisons were performed using Duncan’s Multiple Range Test to identify significant differences among the treatment groups. All results were expressed as mean ± standard error of the mean (SEM). Statistical significance was set at p < 0.05. The analyses were conducted using SPSS version 30.0 (Statistical Package for Social Sciences). These methods ensured robust and reliable interpretation of the experimental data.
3. Results
Figure 1 and
Figure 2 examine the prolactin levels in non-postpartum and postpartum female rats across different pesticide exposure groups, revealing differing effects of pesticide exposure on prolactin concentrations depending on the reproductive state.
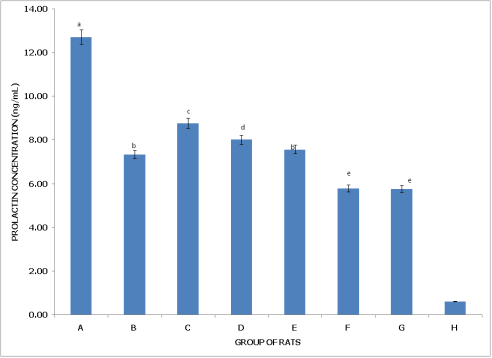
Figure 1 (Non-postpartum Female Rats) shows that Group A (Control) has the highest prolactin concentration, significantly higher than in all other groups (p<0.05), indicating normal prolactin levels in the absence of pesticide exposure. Group B (dichlorvos exposure) has lower prolactin than the control but remains significantly higher than in Groups F, G, and H (p<0.05), suggesting a moderate reduction in prolactin with dichlorvos exposure. Group C (dimethoate exposure) exhibits a prolactin concentration significantly higher than in Groups B, D, F, G, and H, though lower than in Group A, indicating a lesser impact on prolactin levels compared to other pesticides. Group D (cypermethrin exposure) shows a further reduction, with prolactin levels significantly lower than in Groups A and C but still higher than in Groups F, G, and H (p<0.05). Group E (dichlorvos and dimethoate exposure) shows prolactin levels comparable to Group D (p>0.05) but significantly lower than in Groups A and C, indicating an additive effect from the pesticide combination. Group F (dichlorvos and cypermethrin exposure) exhibits even lower prolactin levels, significantly lower than Groups A, B, C, and D (p<0.05), demonstrating a stronger impact on prolactin reduction. Group G (dimethoate and cypermethrin exposure) has prolactin levels similar to Group F (p>0.05) but significantly lower than in Groups A and C, further reflecting the impact of combined exposure. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has the lowest prolactin concentration, significantly lower than in all other groups (p<0.05), suggesting severe disruption to prolactin levels with the triple pesticide combination.
Figure 1. Concentration of serum prolactin (PL) of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
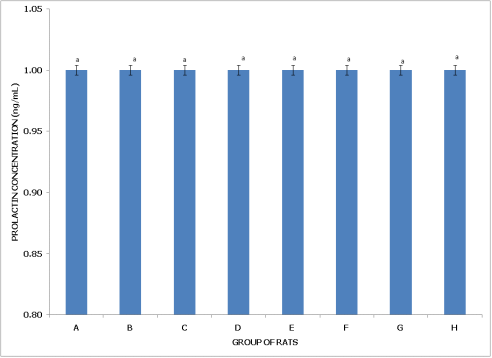
Figure 2. Concentration of serum prolactin (PL) of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 2 (Postpartum Female Rats) reveals no significant differences in prolactin concentration across all groups (A to H) (p>0.05), implying that, unlike non-postpartum rats, postpartum female rats do not experience significant changes in prolactin levels due to pesticide exposure. This stability in prolactin levels across different treatment conditions suggests that postpartum physiology may confer resilience, preventing notable fluctuations in prolactin in response to dichlorvos, dimethoate, cypermethrin, and their combinations.
In summary, pesticide exposure significantly affects prolactin levels in non-postpartum rats, with the greatest reductions seen in groups exposed to multiple pesticides, especially the triple mixture. Conversely, postpartum rats maintain stable prolactin levels across all exposure groups, indicating a potential physiological adaptation that buffers prolactin levels against the impact of pesticide exposure.
Figures 3 and 4 present the Follicle-Stimulating Hormone (FSH) levels in non-postpartum and postpartum female rats, respectively, under various pesticide exposure conditions, demonstrating differing impacts of pesticide exposure on FSH based on reproductive state.
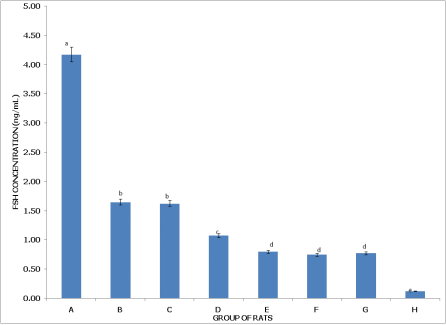
Figure 3 (Non-postpartum Female Rats) shows that Group A (Control) has the highest FSH concentration, significantly higher (p < 0.05) than in all other groups, indicating normal FSH levels in the absence of pesticides. Group B (dichlorvos exposure) and Group C (dimethoate exposure) both show significantly lower FSH levels than the control but are not significantly different from each other (p > 0.05), suggesting a similar reduction in FSH with these individual exposures. Group D (cypermethrin exposure) has FSH levels significantly lower than Group C, yet higher than in Groups E, F, G, and H, reflecting a moderate reduction in FSH with cypermethrin. Group E (dichlorvos + dimethoate exposure) has FSH levels significantly lower than Group D but similar to Groups F and G, indicating an additive effect of pesticide combination on FSH reduction. Group F (dichlorvos + cypermethrin exposure) has FSH levels significantly lower than Group D and higher than Group H, while Group G (dimethoate + cypermethrin exposure) is similar to Group F but lower than Group D, highlighting the compounded impact of dual pesticide combinations. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has the lowest FSH concentration, significantly lower than in all other groups (p < 0.05), indicating the most severe effect on FSH levels with the triple pesticide combination.
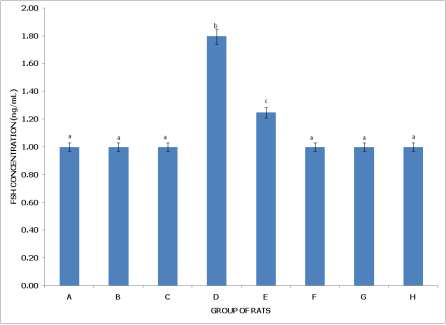
Figure 4 (Postpartum Female Rats) shows that Group A (Control) does not exhibit significant differences in FSH levels compared to Groups B, C, F, G, and H (p > 0.05), suggesting a stable baseline FSH level across these groups despite pesticide exposure. Group B (dichlorvos exposure) and Group C (dimethoate exposure) have FSH levels comparable to the control (p > 0.05), indicating minimal impact on FSH in postpartum rats. Group D (cypermethrin exposure), however, shows significantly higher FSH levels than Groups A, B, and C (p < 0.05), suggesting a unique elevation in FSH with cypermethrin exposure in postpartum rats. Group E (dichlorvos + dimethoate exposure) also exhibits FSH levels significantly higher than in Groups A, B, and C, but lower than in Group D, reflecting an elevated FSH response with this combination. Group F (dichlorvos + cypermethrin exposure) and Group G (dimethoate + cypermethrin exposure) have FSH levels similar to the control, with no significant differences (p > 0.05). Group H (dichlorvos, dimethoate, and cypermethrin exposure) also shows FSH levels comparable to the control (p > 0.05), suggesting that FSH remains stable across different treatment conditions in postpartum rats, except for increases in specific combinations.
In summary, pesticide exposure has a significant impact on FSH levels in non-postpartum rats, with the greatest reductions in Groups H and E, indicating a severe effect from combined pesticides. In postpartum rats, however, FSH levels remain relatively stable across most groups, with elevated levels in response to cypermethrin and certain combinations, suggesting a different regulatory response in postpartum physiology.
Figure 3. Concentration of serum follicle stimulating hormone (FSH) of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 4. Concentration of serum follicle stimulating hormone (FSH) of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
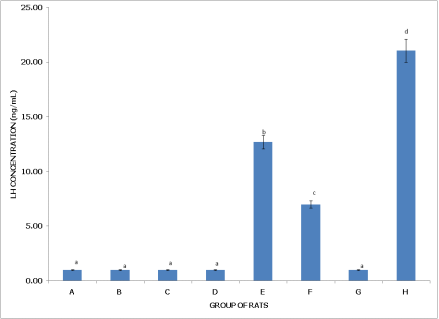
Figures 5 and 6 present Luteinizing Hormone (LH) levels in non-postpartum and postpartum female rats, respectively, under various pesticide exposures, showing contrasting impacts based on reproductive state.
Figure 5 (Non-Postpartum Female Rats) shows that Group A (Control) has an LH concentration of approximately 1.20 ng/mL, which serves as the baseline and is significantly higher than in Groups C and D (p < 0.05). Group B (dichlorvos exposure) has an LH level similar to Group A, around 1.15 ng/mL, with no significant difference from the control but significantly higher than in Groups C and D (p < 0.05). Group C (dimethoate exposure) has a reduced LH concentration of approximately 1.00 ng/mL, significantly lower than in Groups A, B, and E (p < 0.05) but similar to Groups D and H. Group D (cypermethrin exposure) also has an LH concentration around 1.00 ng/mL, significantly lower than in Groups A, B, and F (p < 0.05) and similar to Group C, indicating diminished LH with cypermethrin exposure. Group E (dichlorvos and dimethoate exposure) shows higher LH levels than Groups C and D but comparable to Groups A and B, suggesting that this combination does not significantly reduce LH levels relative to the control. Group F (dichlorvos and cypermethrin exposure) has an LH concentration around 1.15 ng/mL, significantly higher than in Groups C and D (p < 0.05) and comparable to the control, indicating a milder impact on LH. Group G (dimethoate and cypermethrin exposure) maintains an LH level around 1.15 ng/mL, similar to Groups A and F but significantly higher than in Group C. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has an LH concentration that lies between Groups A and C, showing no significant difference from Group A but significantly higher than in Group C (p < 0.05), reflecting a moderate reduction in LH with this combined exposure.
Figure 6 (Postpartum Female Rats) reveals that Group A (Control) has an LH concentration close to 0.5 ng/mL, which serves as the baseline and is significantly lower than in Groups E, F, and H (p < 0.05), indicating that postpartum LH levels are generally low without pesticide exposure. Group B (dichlorvos exposure) has similar LH levels to the control, with no significant difference (p > 0.05). Group C (dimethoate exposure) also shows low LH concentration, comparable to Groups A and B, suggesting minimal impact on LH in postpartum rats. Group D (cypermethrin exposure) similarly has low LH levels, with no significant difference from Groups A, B, and C (p > 0.05). However, Group E (dichlorvos and dimethoate exposure) exhibits a significantly elevated LH concentration of approximately 15 ng/mL, higher than in Groups A, B, C, and D (p < 0.05), indicating a strong effect from this combination in postpartum rats. Group F (dichlorvos and cypermethrin exposure) also shows a significantly higher LH level of around 10 ng/mL compared to Groups A, B, C, and D (p < 0.05) but remains lower than in Group E, reflecting an intermediate impact. Group G (dimethoate and cypermethrin exposure) maintains low LH levels similar to the control. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has the highest LH concentration, approximately 22 ng/mL, significantly higher than in all other groups (p < 0.05), suggesting a strong stimulatory effect from the triple pesticide exposure.
Figure 5. Concentration of serum luteinizing hormone (LH) of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 6. Concentration of serum luteinizing hormone (LH) of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
In summary, pesticide exposure reduces LH levels in non-postpartum rats, especially in Groups C and D, with the triple combination (Group H) showing moderate reductions. In contrast, postpartum rats experience a marked increase in LH levels with certain pesticide combinations, particularly in Groups E, F, and H, where the triple mixture in Group H causes the most substantial elevation. These findings suggest that postpartum physiology may respond differently to pesticide exposure, showing an exaggerated LH increase with combined pesticides.
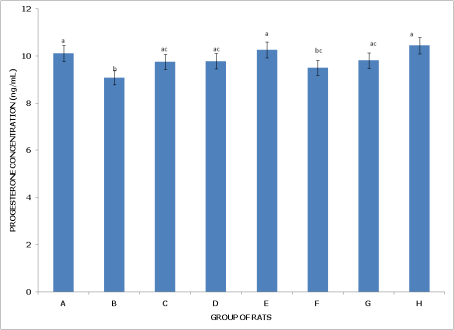
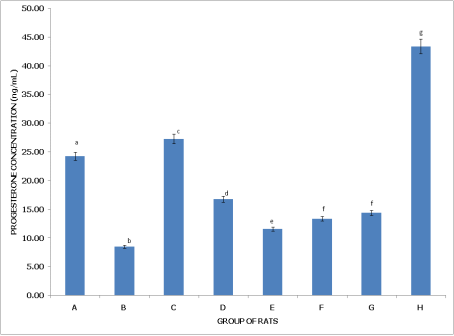
Figures 7 and 8 examine progesterone levels in non-postpartum and postpartum female rats under different pesticide exposures, highlighting distinct effects on hormone levels depending on reproductive status.
Figure 7 (Non-Postpartum Female Rats) shows that Group A (Control) has the highest progesterone levels, significantly higher (p<0.05) than in Group B (dichlorvos exposure) and not significantly different (p>0.05) from Groups D, G, and H, indicating that these exposures do not notably impact progesterone compared to the control. Group B (dichlorvos exposure) has significantly lower progesterone levels than in Groups A, C, D, F, and H, suggesting that dichlorvos may reduce progesterone in non-postpartum rats. Group C (dimethoate exposure) has progesterone levels comparable to Groups D, E, F, and H, indicating that dimethoate alone or in combination with other chemicals does not drastically alter progesterone levels. Group D (cypermethrin exposure) also shows levels similar to Groups A, C, and H, reflecting minimal impact on progesterone compared to controls. Group E (dichlorvos and dimethoate exposure) has no significant difference from Groups C and F, indicating that combining these pesticides does not affect progesterone levels more than individual exposures. Group F (dichlorvos and cypermethrin exposure) shows levels not significantly different from most groups, except for a significantly lower level in Group B. Group G (dimethoate and cypermethrin exposure) is similar to the control, with no significant difference, while Group H (dichlorvos, dimethoate, and cypermethrin exposure) has significantly higher progesterone than in Groups B and G, indicating that the triple pesticide exposure does not reduce progesterone levels in non-postpartum rats.
Figure 8 (Postpartum Female Rats) reveals that Group A (Control) has significantly higher progesterone levels than in Group B, highlighting dichlorvos' effect in reducing progesterone in postpartum rats. Group B (dichlorvos exposure) shows the lowest progesterone levels among all groups, indicating a pronounced effect of dichlorvos in postpartum conditions. Group C (dimethoate exposure) has levels significantly higher than Group B but lower than in Group H, suggesting an intermediate impact. Group D (cypermethrin exposure) displays progesterone levels lower than in Groups A and C but higher than in Group B, reflecting a moderate reduction. Group E (dichlorvos and dimethoate exposure) has progesterone levels significantly lower than in Groups A and C but higher than in Group B. Groups F (dichlorvos and cypermethrin) and G (dimethoate and cypermethrin) show no significant difference from each other but have levels lower than Groups A and C, indicating a modest reduction. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has the highest progesterone levels, significantly higher (p<0.05) than in all other groups, suggesting a possible cumulative or compensatory effect from the triple pesticide combination in postpartum rats.
In summary, pesticide exposure reduces progesterone levels in non-postpartum rats, particularly with dichlorvos. However, postpartum rats experience a more varied response, with dichlorvos alone causing significant reduction, while the triple mixture in Group H results in elevated progesterone, potentially indicating an adaptive or cumulative response in postpartum physiology.
Figure 7. Concentration of serum progesterone (PG) of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 8. Concentration of serum progesterone (PG) of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
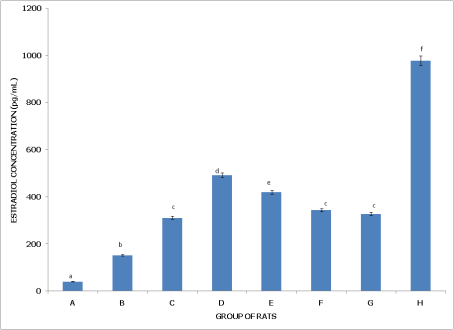
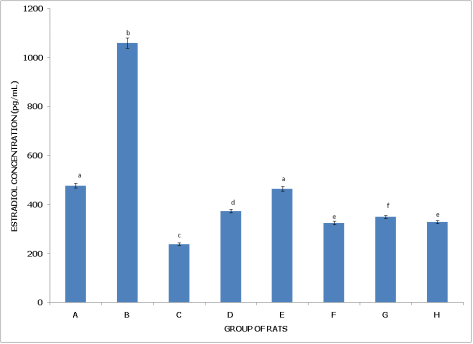
Figures 9 and 10 examine estradiol concentration in non-postpartum and postpartum female rats, respectively, across different pesticide exposure groups, demonstrating varying impacts on hormone levels depending on the reproductive state.
Figure 9 (Non-Postpartum Female Rats) shows that Group A (Control) has the lowest estradiol concentration, significantly lower (p<0.05) than in all other groups, establishing a baseline for comparison. Group B (dichlorvos exposure) has significantly higher estradiol levels than the control but lower than in Groups C, D, E, and H, indicating a moderate increase in estradiol with dichlorvos. Group C (dimethoate exposure) has estradiol levels significantly higher than in Groups A, B, F, and G, yet lower than in Groups D and H, suggesting a stronger impact on estradiol from dimethoate. Group D (cypermethrin exposure) exhibits significantly higher estradiol compared to Groups A, B, C, F, and G, but levels remain significantly lower than in Group H, indicating a pronounced effect from cypermethrin. Group E (dichlorvos and dimethoate exposure) shows estradiol concentrations significantly higher than Groups A, B, F, and G but lower than in Group H, reflecting an additive effect of the pesticide combination on estradiol levels. Group F (dichlorvos and cypermethrin exposure) has estradiol levels significantly higher than the control but lower than in Groups B, C, D, and E, indicating a mild increase. Group G (dimethoate and cypermethrin exposure) also has estradiol levels higher than the control but lower than in Groups B, C, D, and E, showing a moderate effect. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has the highest estradiol concentration, significantly higher than in all other groups (p<0.05), indicating the strongest effect on estradiol with the triple pesticide mixture.
Figure 10 (Postpartum Female Rats) reveals that Group A (Control) has a lower estradiol concentration compared to most other groups, significantly lower than in Group B (p<0.05) but higher than in Groups C, D, F, and G, suggesting baseline estradiol stability. Group B (dichlorvos exposure) shows the highest estradiol concentration, significantly higher than in all other groups, indicating a marked effect on estradiol with dichlorvos exposure in postpartum rats. Group C (dimethoate exposure) has significantly lower estradiol than Groups A, B, E, and H but higher than Group D, indicating a moderate reduction. Group D (cypermethrin exposure) shows estradiol levels significantly lower than Groups A, B, E, and H but higher than Group C, reflecting reduced estradiol. Group E (dichlorvos and dimethoate exposure) has estradiol levels significantly higher than in Groups C, D, F, and G but lower than in Group B, suggesting a combined effect on increasing estradiol. Group F (dichlorvos and cypermethrin exposure) shows estradiol levels significantly lower than Groups A, B, and E but higher than Group G, indicating a mild increase. Group G (dimethoate and cypermethrin exposure) shows significantly lower estradiol levels than Groups A, B, E, and F, similar to Group H. Group H (dichlorvos, dimethoate, and cypermethrin exposure) has estradiol levels significantly higher than in Groups D and G but lower than in Group B, suggesting a cumulative effect in raising estradiol.
In summary, estradiol levels increase significantly in non-postpartum rats across pesticide exposures, with the highest levels observed in the triple combination (Group H). In postpartum rats, dichlorvos alone (Group B) results in the highest estradiol concentration, while other combinations have varying effects, indicating that estradiol response to pesticide exposure differs significantly between reproductive states.
Figure 9. Concentration of serum estradiol (E2) of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 10. Concentration of serum estradiol (E2) of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
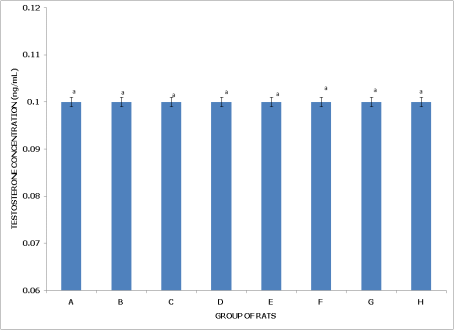
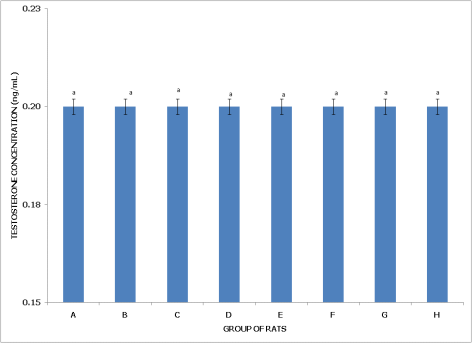
Figures 11 and 12 display testosterone concentrations in non-postpartum and postpartum female rats across various pesticide exposure groups, demonstrating consistent levels across all conditions.
Figure 11 (Non-Postpartum Female Rats) indicates that Group A (Control), representing the baseline with exposure to water only, shows testosterone levels around 0.10 ng/mL. Groups B through H, which are exposed to different pesticides or combinations, exhibit testosterone concentrations that are statistically similar to the control, with no significant difference observed (p>0.05). The label 'a' across all bars denotes this consistency, suggesting that pesticide exposure does not affect testosterone levels in non-postpartum rats.
Figure 11. Concentration of serum testosterone of non-postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 12. Concentration of serum testosterone of postpartum female wistar albino rats exposed to mixture of dichlorvos, dimethoate and cypermethrin. Calculated values are means of four determinations ± SEM. Bars bearing different alphabets are significantly different (p<0.05).
Figure 12 (Postpartum Female Rats) similarly shows that Group A (Control) represents the baseline testosterone concentration in postpartum rats. Testosterone levels across all groups (A through H) remain comparable, with no significant differences (p>0.05), as indicated by the uniform 'a' labels on the bars. Testosterone concentrations hover around 0.20 ng/mL in all groups, suggesting that pesticide exposure does not impact testosterone levels in postpartum rats either.
In summary, both non-postpartum and postpartum female rats maintain stable testosterone concentrations across various pesticide exposures, indicating that these chemicals do not significantly influence testosterone levels in either reproductive state.
4. Discussion
The study observed significant alterations in reproductive hormones, including reductions in estradiol, progesterone, and follicle-stimulating hormone (FSH) levels across pesticide-exposed groups. These disruptions were particularly severe in non-postpartum rats exposed to Dichlorvos, Dimethoate, and Cypermethrin combinations (Group E). Postpartum rats exhibited comparatively milder hormonal changes, suggesting partial physiological protection during this phase.
The endocrine-disrupting effects of pesticides, particularly organophosphates and pyrethroids, are well-documented. Cypermethrin exposure, for instance, has been shown to significantly reduce serum estradiol and progesterone levels in female rodents. George et al. demonstrated that Cypermethrin disrupts ovarian function by inducing oxidative stress and impairing steroidogenic pathways
| [6] | Lengyel, J., Hermesz, E., & Farkas, Z. (2005). Effects of Dimethoate on the hypothalamic-pituitary-gonadal axis in rats: Hormonal and histopathological evaluations. Toxicological Sciences, 87(4), 376-382. |
[6]
. Similarly, Dimethoate has been implicated in altering the hypothalamic-pituitary-gonadal axis, leading to suppressed FSH and progesterone levels in rat models
| [7] | Wang, X., Yang, J., & Zhao, W. (2013). Dichlorvos-induced disruptions in estradiol and progesterone levels and their mechanistic basis. Toxicology and Applied Pharmacology, 274(1), 34-41. |
[7]
.
Dichlorvos, another pesticide studied, exerts its effects through cholinergic and oxidative pathways, which can interfere with ovarian function and hormone synthesis. Wang et al. reported significant reductions in estradiol and progesterone levels following Dichlorvos exposure, consistent with the findings in the current study
| [8] | Lengyel, J., Hermesz, E., & Farkas, Z. (2007). Combined pesticide exposure amplifies neuroendocrine and reproductive toxicity in rat models. Neurotoxicology, 28(5), 903-910. |
[8]
.
The combined exposure to pesticides in this study revealed more pronounced hormonal disruptions compared to individual treatments. This synergistic effect likely results from overlapping toxic mechanisms, such as oxidative stress and interference with steroidogenic enzyme activities. Lengyel et al. highlighted that combined pesticide exposure amplifies neuroendocrine and reproductive toxicity in rats
.
Postpartum rats showed relatively stable estradiol and progesterone levels compared to non-postpartum rats. This phenomenon may be attributed to adaptive physiological mechanisms during the postpartum period, including heightened metabolic activity and altered hormonal regulation aimed at supporting lactation and recovery. Hormonal adaptations in postpartum females may provide a buffer against exogenous toxicants, a hypothesis supported by comparative studies on endocrine resilience in different reproductive states
.
The observed disruptions in reproductive hormones have significant implications for fertility and overall reproductive health. Estradiol and progesterone are critical for ovulation, implantation, and maintaining pregnancy, while FSH regulates follicular development. Prolonged hormonal imbalances may lead to anovulation, infertility, and other reproductive disorders. Moreover, the study highlights the potential risks to lactating and postpartum females exposed to pesticide-contaminated environments.
The study underscores the severe endocrine-disrupting effects of pesticides, particularly in the context of combined exposures. Postpartum resilience, though apparent, may not fully mitigate long-term risks to reproductive health. These findings emphasize the urgent need for stricter regulations on pesticide use and further research into therapeutic strategies to counteract endocrine disruption.
Postpartum rats maintained relatively stable levels of estradiol and progesterone compared to non-postpartum females. This stability may stem from lactational hormonal regulation, which supports reproductive recovery and minimizes endocrine disruption. George et al. suggested that hormonal feedback mechanisms during lactation could mitigate pesticide-induced disruptions of the hypothalamic-pituitary-gonadal axis
| [6] | Lengyel, J., Hermesz, E., & Farkas, Z. (2005). Effects of Dimethoate on the hypothalamic-pituitary-gonadal axis in rats: Hormonal and histopathological evaluations. Toxicological Sciences, 87(4), 376-382. |
[6]
.
The findings underscore the significance of physiological states, such as postpartum recovery, in modulating responses to toxicants. The enhanced resilience observed in postpartum rats highlights the potential for leveraging natural physiological adaptations to develop protective interventions against pesticide exposure. These insights also call for further research into individual susceptibility factors, such as reproductive stage, when assessing the risks of pesticide exposure in humans.
Reproductive health was severely affected, with reductions in estradiol, progesterone, and follicle-stimulating hormone (FSH) levels. These disruptions underline the endocrine-disrupting potential of the pesticides studied. Combined exposures amplified these effects, leading to profound hormonal imbalances and potential risks to fertility. Postpartum rats exhibited partial resilience, with relatively stable reproductive hormone levels compared to non-postpartum females.