A survey was undertaken from March to June 2014 on the biodiversity and the community structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River mouths (South-Cameroon). In each river, raw waters were collected from upstream to downstream at four sites. Cells were counted using the Malassez cells procedure and species were identified. A total of 10427.1x105 cells corresponded to three phyla, eight classes, 23 orders, 32 genera and 40 species (24 freshwater species (60.0% of total species richness and total collection respectively), three marine species (7.5% and 2.4% of the total species richness; and total collection respectively), and one brackish water specialist in Kienke (2.5% and 5.1%), 13 tolerant species (32.5% and 32.6%)). The trophic diatom index revealed undisturbed conditions with no or little alteration of human origin and a low organic pollution (oligotrophic or mesotrophic state) (Nyong: TDI=52.7; Kienke: TDI=69.7; pooled assemblage: TDI=65.0). A low species richness was detected (richness ratio in Nyong: d=0.008; Kienke: d=0.003; pooled rivers: d=0.004), a high species diversity (Shannon index close to maximum) (Nyong: H’=2.742 and H’max=2.996; Kienke: H’=2.685 and H’max=2.996; pooled rivers: H’=3.245 and H’max=3.689), a very low dominance by a few species (Berger-Parker index close to 0) (Nyong: IBP=0.156; Kienke: IBP=0.175; pooled rivers: IBP=0.134), and Hill’s ratio were close to 1 (Nyong: Hill=0.819; Kienke: Hill=0.803; pooled rivers: Hill=0.722). The community was highly even with a high value of the Pielou’s evenness close to 1 (Nyong: J=0.915; Kienke: J=0.896; pooled rivers: J=0.880). Two useful species and one harmful species to fish were rare in Kienke. Species exhibited in Kienke and pooled data in rainy season, a positive global net association while it was negative in Nyong. Assemblage fitted Preston’s model in Nyong with a high environmental constant in the dry season (m’=1.469), low constant in the rainy season (m’=0.947) and the pooled seasons (m’=0.853). In Kienke constants were low (dry season: m’=0.574; rainy season: m’=0.566; pooled seasons: m’=0.581) suggesting a evolved community in less disturbed environments where the majority of species showed moderate abundances. In the dry season, the pooled assemblage functionned on the basis of maintaining a complex information network (close to ecological balance) developed at spatio-temporal scales (ZM model) and it presented a low force of regeneration (fractal dimension of the distribution of individuals among species (1/γ)=0.925<1). The evolved oligotrophic state (close to natural balance) of the chromists’ community should be preserved and protected and the studied rivers classified as reference.
| Published in | International Journal of Ecotoxicology and Ecobiology (Volume 9, Issue 1) |
| DOI | 10.11648/j.ijee.20240901.12 |
| Page(s) | 28-55 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Freshwater Species, Microalgae, Species Composition, Useful Species, Assemblage Functioning, Water Quality
Families/Species | References | A1 (%) | A2 (%) | Total (%) | B1 (%) | B2 (%) | Total (%) | |
|---|---|---|---|---|---|---|---|---|
Achnanthaceae Kütz., 1844 | ||||||||
Ac. exiguoides#,BI | [43] | - | 125.0 (1.2) | 125.0 (1.2) | - | - | - | |
Amphipleuraceae Grunow (d), 1862 | ||||||||
Fr. adnata #,BI | [49] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Anomoeoneidaceae D. G. Mann, 1990 | ||||||||
An. sphaerophora*,#,BI | [21, 43] | - | 125.0 (1.2) | 125.0 (1.2) | - | - | - | |
Aulacoseiraceae R. M. Crawford, 1990 | ||||||||
Au. granulata #,EI | [13, 19-21, 48, 49] | - | 229.2 (2.2) | 229.2 (2.2) | - | - | - | |
Bacillariaceae Ehrenb., 1831 | ||||||||
De. elegans #,BI | [11] | 62.5 (0.6) | - | 62.5 (0.6) | - | - | - | |
De. thermalis *,BI | [11] | - | - | - | - | 531.3 (5.1) | 531.3 (5.1) | |
Ha. amphioxys †,#,‡,BI | [21, 43, 49] | - | 218.8 (2.1) | 218.8 (2.1) | - | - | - | |
Ni. amphibia #,BI | [16, 22, 43, 48, 49] | - | - | - | - | 375.0 (3.6) | 375.0 (3.6) | |
Ni. sigma #,BI | [22, 43, 49] | - | 20.8 (0.2) | 20.8 (0.2) | - | - | - | |
Ni. tryblionella #,†,BI | [43] | - | - | - | - | 62.5 (0.6) | 62.5 (0.6) | |
Catenulaceae Mereschkowsky, 1902 | ||||||||
Amphora ovalis #,BI | [21, 43] | - | 291.7 (2.8) | 291.7 (2.8) | - | - | - | |
Ceratiaceae Kofoid, 1907 | ||||||||
Ce. hirundinella #,†,BL | [44] | - | - | - | - | 312.5 (3.0) | 312.5 (3.0) | |
Chaetocerotaceae Ralfs (d), 1861 | ||||||||
Ch. muelleri †,US,ITP | [21] | - | - | - | 62.5 (0.6) | - | 62.5 (0.6) | |
Cocconeidaceae Kützing, 1844 | ||||||||
Co. placentula #,†,BI | [14, 21, 43, 48] | - | 145.8 (1.4) | 145.8 (1.4) | - | - | - | |
Coscinodiscaceae Kützing, 1844 | ||||||||
Cs. rudolfi †,BI | [15, 43] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Cryptomonadaceae Ehrenberg, 1831 | ||||||||
Cr. erosa *,#,BI | [52] | - | - | - | 375.0 (3.5) | - | 375.0 (3.5) | |
Dinobryaceae Ehrenberg, 1834 | ||||||||
Di. sertularia *,#,BI | [52] | - | - | - | - | 83.3 (0.8) | 83.3 (0.8) | |
Diploneidaceae D. G. Mann (d), 1990 | ||||||||
Dp. arctica #,BI | [63] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Dp. ovalis #,BI | [11] | - | - | - | - | 145.8 (1.4) | 145.8 (1.4) | |
Fragilariaceae Kützing, 1844 | ||||||||
Fr. construens *,#,BI | [13, 21, 43] | - | 125.0 (1.2) | 125.0 (1.2) | - | - | - | |
Synedra ulna #,BI | [16, 43, 49] | - | - | - | - | 531.3 (5.1) | 531.3 (5.1) | |
Gomphonemataceae Kützing, 1844 | ||||||||
Go. olivaceum #,BI | [16, 48] | - | - | - | - | 1000.0 (9.6) | 1000.0 (9.6) | |
Goniochloridaceae Bailey (d) | ||||||||
Gn. gigas #,BI | [43] | - | - | - | - | 666.7 (6.4) | 666.7 (6.4) | |
Gn. mutica #,BI | [45] | - | 83.3 (0.8) | 83.3 (0.8) | - | - | - | |
Mastogloiaceae Mereschkowsky, 1903 | ||||||||
Ma. smithii #,BI | [43] | - | - | - | - | 62.5 (0.6) | 62.5 (0.6) | |
Melosiraceae Kützing, 1844 | ||||||||
Me. granulata #,†,BI | [43, 48] | - | - | - | - | 1395.8 (13.4) | 1395.8 (13.4) | |
Ophiocytiaceae Lemmermann, 1899 | ||||||||
Op. cochleare #,BI | [50] | 20.8 (0.2) | - | 20.8 (0.2) | - | - | - | |
Phytodiniaceae Klebs, 1912 | ||||||||
Cy. unicorne #,BI | [44] | - | - | - | - | 281.3 (2.6) | 281.3 (2.6) | |
Pinnulariaceae D. G. Mann (d), 1990 | ||||||||
Pi. cardinaliculus #,BI | [62] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Rhizosoleniaceae De Toni, 1890 | ||||||||
Rh. longiseta #,BI | [43] | - | - | - | - | 729.2 (7.0) | 729.2 (7.0) | |
Rhopalodiaceae Topachevs'kyj (d) & Oksiyuk (d), 1960 | ||||||||
Ep. turgida †,US(NF) | [16, 23, 48] | - | - | - | - | 125.0 (1.2) | 125.0 (1.2) | |
Stephanodiscaceae I. V. Makarova (d), 1986 | ||||||||
Cc. meneghiniana #,†,BI | [43, 48, 49] | 62.5 (0.6) | - | 62.5 (0.6) | - | - | - | |
Cc. stelligera #,BI | [17, 43, 48 49] | - | - | - | - | 333.3 (3.2) | 333.3 (3.2) | |
St. astraea #,†,BI | [13] | - | - | - | - | 406.3(3.9) | 406.3(3.9) | |
Surirellaceae Kützing, 1844 | ||||||||
Ca. noricus #,BI | [11] | - | 385.4 (3.7) | 385.4 (3.7) | - | - | - | |
Cm. apiculata #,BI | [21] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Cm. solea #,BI | [21] | - | 187.5 (1.8) | 187.5 (1.8) | - | - | - | |
Su. capronii #,BI | [43] | - | - | - | - | 239.6 (2.3) | 239.6 (2.3) | |
Su. linearis #,BI | [43] | - | - | - | 250.0(2.3) | - | 250.0 (2.3) | |
Tabellariaceae Kützing, 1844 | ||||||||
Ta. flocculosa #,‡,BI | [13, 22, 43, 48, 49] | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - | |
Total | 145.8(1.4) | 2312.5(22.2) | 2458.3(23.6) | 687.5(6.6) | 7281.3(69.8) | 7968.8(76.4) | ||
Families | Species | References | C1 (%) | C2 (%) | Total (%) |
|---|---|---|---|---|---|
Achnanthaceae | Achnanthes exiguoides #,BI | [43] | - | 125.0 (1.2) | 125.0 (1.2) |
Amphipleuraceae | Frustulia adnata #,BI | [49] | - | 62.5 (0.6) | 62.5 (0.6) |
Anomoeoneidaceae | Anomoeoneis sphaerophora *,#,BI | [21] | - | 125.0 (1.2) | 125.0 (1.2) |
Aulacoseiraceae | Aulacoseira granulata #,EI, | [13, 19-21, 48, 49] | - | 229.2 (2.2) | 229.2 (2.2) |
Bacillariaceae | Denticula elegans #,BI | [11] | 62.5 (0.6) | - | 62.5 (0.6) |
De. thermalis var. fossilis *,BI | [11] | - | 531.3 (5.1) | 531.3 (5.1) | |
Hantzschia amphioxys †,#,‡,BI | [21, 43, 49] | - | 218.8 (2.1) | 218.8 (2.1) | |
Nitzschia amphibia #,BI | [16, 18, 21, 43, 48, 49] | - | 375.0 (3.6) | 375.0 (3.6) | |
Ni. sigma #,BI | [21, 43, 49] | - | 20.8 (0.2) | 20.8 (0.2) | |
Ni. tryblionella #,†,BI | [43] | - | 62.5 (0.6) | 62.5 (0.6) | |
Catenulaceae | Amphora ovalis #,BI | [21, 43] | - | 291.7 (2.8) | 291.7 (2.8) |
Ceratiaceae | Ceratium hirundinella #,†,BI | [44] | - | 312.5 (3.0) | 312.5 (3.0) |
Chaetocerotaceae | Chaetoceros muelleri †,ITP | [21] | 62.5 (0.6) | - | 62.5 (0.6) |
Cocconeidaceae | Cocconeis placentula #,†,BI | [14, 21, 43, 48] | - | 145.8 (1.4) | 145.8 (1.4) |
Coscinodiscaceae | Coscinodiscus rudolfi †,BI | [15, 43] | - | 62.5 (0.6) | 62.5 (0.6) |
Cryptomonadaceae | Cryptomonas erosa *,#,BI | [52] | 375.0 (3.6) | - | 375.0 (3.6) |
Dinobryaceae | Dinobryon sertularia *,#,BI | [52] | - | 83.3 (0.8) | 83.3 (0.8) |
Diploneidaceae | Diploneis arctica #,BI | [63] | - | 62.5 (0.6) | 62.5 (0.6) |
Dp. ovalis var. pumila #,BI | [11] | - | 145.8 (1.4) | 145.8 (1.4) | |
Fragilariaceae | Fragilaria construens *,#,BI | [13, 16, 21, 43] | - | 125.0 (1.2) | 125.0 (1.2) |
Synedra ulna #, BI | [43, 49] | - | 531.3 (5.1) | 531.3 (5.1) | |
Gomphonemataceae | Gomphonema olivaceum #,BI | [16, 48] | - | 1000.0 (9.6) | 1000.0 (9.6) |
Goniochloridaceae | Goniochloris gigas #,BI | [43] | - | 666.7 (6.4) | 666.7 (6.4) |
Gn. mutica #,UN(BI) | [45] | - | 83.3 (0.8) | 83.3 (0.8) | |
Mastogloiaceae | Mastogloia smithii #,BI | [43] | - | 62.5 (0.6) | 62.5 (0.6) |
Melosiraceae | Melosira granulata #,†,BI | [43, 48] | - | 1395.8(13.4) | 1395.8 (13.4) |
Ophiocytiaceae | Ophiocytium cochleare #,BI | [50] | 20.8 (0.2) | - | 20.8 (0.2) |
Phytodiniaceae | Cystodinium unicorne #,BI | [44] | - | 281.3 (2.6) | 281.3 (2.6) |
Pinnulariaceae | Pinnularia cardinaliculus #,BI | [62] | - | 62.5 (0.6) | 62.5 (0.6) |
Rhizosoleniaceae | Rhizosolenia longiseta #,BI | [43] | - | 729.2 (7.0) | 729.2 (7.0) |
Rhopalodiaceae | Epithemia turgida †, US(BF) | [16, 23, 48] | - | 125.0 (1.2) | 125.0 (1.2) |
Stephanodiscaceae | Cyclotella meneghiniana #,†,BI | [21, 43, 48, 49] | 62.5 (0.6) | - | 62.5 (0.6) |
Cc. stelligera #, BI) | [17, 43, 48, 49] | - | 333.3 (3.2) | 333.3 (3.2) | |
Stephanodiscus astraea #,†,BI | [13] | - | 406.3 (3.9) | 406.3 (3.9) | |
Surirellaceae | Campylodiscus noricus #,BI | [11] | - | 385.4 (3.7) | 385.4 (3.7) |
Cm. apiculata #,BI | [21] | - | 62.5 (0.6) | 62.5 (0.6) | |
Cm. solea #,BI | [21] | - | 187.5 (1.8) | 187.5 (1.8) | |
Surirella capronii #,BI | [43] | - | 239.6 (2.3) | 239.6 (2.3) | |
Su. linearis #,BI | [43] | 250.0 (2.4) | - | 250.0 (2.4) | |
Tabellariaceae | Ta. flocculosa #,‡,,BI | [13, 16, 21, 22, 43, 48, 49] | - | 62.5 (0.6) | 62.5 (0.6) |
Total | 833.3 8.0) | 9593.8 92.0) | 10427.1(100.0) |
Water | Nyong River mouth | Kienke River mouth | ||||
|---|---|---|---|---|---|---|
A. (%) | B (%) | Pooled (%) | A (%) | B (%) | Pooled rivers (%) | |
Abundance of the specialist species x105 (%) | ||||||
Freshwater | 62.5 (0.6) | 1572.9 (15.1) | 1635.4 (15.7) | 250.0 (2.4) | 4364.6 (41.9) | 4614.6 (44.3) |
Brackish | - | - | - | - | 531.3 (5.1) | 531.3 (5.1) |
Marine | - | 62.5 (0.6) | 62.5 (0.6) | 62.5 (0.6) | 125.0 (1.2) | 187.5 (1.8) |
Total 1 | 62.5 (0.6) | 1635.4 (15.7) | 1697.9 (16.3) | 312.5 (3.0) | 5060.8 (48.5) | 5333.3 (51.1) |
Test (FE or FFH): | - | p<0.001* | p<0.001* | p<0.001* | FFH: df=2, p<0.001 * | FFH: df=2, p<0.001 * |
Avs.B: FE (df=1) | I: p<0.001*; III: p-2.0x10-19 *; Total: p<0.001* | I: p<0.001*; II: p<0.001*; III: p=6.6x10-6 *; Total: FE: p<0.001* | ||||
Abundance of the tolerant species x105 (%) | ||||||
I | 83.3 (0.8) | 145.8 (1.4) | 229.2 (2.2) | - | 2177.1(20.9) | 2177.1 (20.9) |
II | - | - | - | 375.0 (3.6) | 83.3 (0.8) | 458.3 (4.4) |
III | - | 62.5 (0.6) | 62.5 (0.6) | - | - | - |
IV | - | 250.0 (2.4) | 250.0 (2.4) | - | - | - |
V | - | 218.8 (2.1) | 218.8 (2.1) | - | - | - |
Total 2 | 83.3 (0.8) | 677.1 (6.5) | 760.4 (7.3) | 375.0 (3.6) | 2260.4 (21.7) | 2635.4 (25.3) |
Global. | 145.8 (1.4) | 2312.5 (22.2) | 2458.3 (23.6) | 687.5 (6.6) | 7281.3 (69.8) | 7968.8 (76.4) |
Total 1vs.Total 2 | p=0.114 ns | p=.5x10-101 * | p<0.001* | p=0.018 * | p<0.001* | p<0.001* |
Test | - | FFH: df=3, p<0.001 * | FFH: df=3 p<0.001 * | - | FE: df=1, p<0.001* | FE: df=1, p<0.001* |
Total 1 vs. Total 2 | p<0.001* | p<0.001* | p<0.001* | p=0.018 ns | p<0.001* | p<0.001* |
Avs.B (FE) | IV: df=1, p<0.001*; VI: df=1, p<0.001* VII: df=1, p<0.001*; VIII: df=1, p<0.001* Total: df=1, p<0.001* | IV: df=1, p<0.001*; V: df=1, p<0.001*; Total: df=1, p<0.001* | ||||
Water | Both River mouths | ||
|---|---|---|---|
A. (%) | B (%) | Pooled rivers (%) | |
Abundance of the specialist species x105 (%) | |||
Freshwater | 312.5 (3.0) | 5937.5 (56.9) | 6250.0 (59.9) |
Brackish | - | 531.3 (5.1) | 531.3 (5.1) |
Marine | 62.5 (0.6) | 187.5 (1.8) | 250.0 (2.4) |
Total 1 | 375.0 (3.6) | 6656.3 (63.8) | 7031.3 (67.4) |
Test: | FE: df=1, p<0.001 * | FFH: df=2, p<0.001 * | FFH: df=2, p<0.001 * |
Avs.B: FE (df=1) | I: p<0.001*; II. p<0.001*; III; p=6.6x10-6 *; Total: FE: p<0.001* | ||
Abundance of the tolerant species x105 (%) | |||
I | 83.3 (0.8) | 2322.9 (22.3) | 2406.3(23.1) |
II | 375 (3.6) | 83.3 (0.8) | 458.3 (4.4) |
III | - | 62.5 (0.6) | 62.5 (0.6) |
IV | - | 250.0 (2.4) | 250.0 (2.4) |
V | - | 218.8 (2.1) | 218.8 (2.1) |
Total 2 | 458.3 (4.4) | 2937.5 (28.2) | 3395.8 (32.6) |
Global. | 833.3 (8.0) | 9593.8 (92.0) | 10427.1 (100.0) |
Test | FE: df=1, p<0.001* | FFH: df=4, p<0.001 * | FFH: df=4, p<0.001 * |
Avs.B (FE) | IV: df=1, p<0.001*; V: df=1, p<0.001*; VI: df=1, p<0.001*; VII: df=1, p<0.001*; VIII: df=1, p<0.001*; Total: df=1, p<0.001* | ||
Comparison: Nyong River mouth vs. Kienke River mouth (Fisher’s exact test) | |||
Freshwater (df=1) | p<0.001* | p<0.001* | p<0.001* |
Brackish | - | p<0.001* | p<0.001* |
Marine | p<0.001* | p=6.6x10-6 * | p=8.6x10-16 * |
Total 1 | p<0.001* | p<0.001* | p<0.001* |
I | p<0.001* | p<0.001* | p<0.001* |
II | - | p<0.001* | p<0.001* |
III | - | p<0.001* | p<0.001* |
IV | - | p<0.001* | p<0.001* |
V | - | p<0.001* | p<0.001* |
Total 2 | p<0.001* | p<0.001* | p<0.001* |
Global | p<0.001* | p<0.001* | p<0.001* |
Total 1 vs. Total 2 | p=3.7x10-3 * | p<0.001* | p<0.001* |
Indices | Nyong River mouths | Kienke River mouths | Both rivers | ||||||
|---|---|---|---|---|---|---|---|---|---|
I | II | III | I | II | III | I | II | III | |
A. Richness indexes | |||||||||
n x105 cells | 145.8 | 2,312.5 | 2,458.3 | 687.5 | 7,281.3 | 7,968.8 | 833.3 | 9,593.8 | 10,427.1 |
(%) | (1.4) | (22.2) | (23.6) | (6.6) | (69.8) | (76.4) | (8.0) | (92.0) | (100.0) |
S (%) | 3 (7.5) | 17 (42.5) | 20 (50.0) | 3 (7.5) | 17 (42.5) | 20 (50.0) | 6 (15.0) | 34 (85.0) | 40 (100.0) |
nmaxx105 cells | 62.5 | 385.4 | 385.4 | 375.0 | 1,395.8 | 1,395.8 | 375.0 | 1,395.8 | 1,395.8 |
Magalef: Mg | 0.401 | 2.065 | 2.433 | 0.306 | 1.799 | 2.115 | 0.743 | 3.599 | 4.215 |
Richness ratio: d = S/n | 0.020 | 0.007 | 0.008 | 0.004 | 0.002 | 0.003 | 0.007 | 0.004 | 0.004 |
Chao1 | 3 | 17 | 20 | 3 | 17 | 20 | 6 | 34 | 40 |
% SE=(S/Chao 1)*100 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
B. Diversity indexes | |||||||||
Shannon-Weaver H’ | 1.004 | 2.612 | 2.742 | 0.918 | 2.530 | 2.685 | 1.398 | 3.102 | 3.245 |
H’max=ln(S) | 1.099 | 2.833 | 2.996 | 1.099 | 2.833 | 2.996 | 1.792 | 3.526 | 3.689 |
Simpson’s D | 0.388 | 0.087 | 0.079 | 0.438 | 0.098 | 0.085 | 0.309 | 0.061 | 0.054 |
Hill’s N1 = eH’ | 2.730 | 13.629 | 15.522 | 2.503 | 12.553 | 14.654 | 4.048 | 22.250 | 25.656 |
Hill’s N2 = 1/D | 2.579 | 11.447 | 12.719 | 2.286 | 10.213 | 11.766 | 3.236 | 16.276 | 18.525 |
Hill’s ratio N2/N1 | 0.945 | 0.840 | 0.819 | 0.913 | 0.814 | 0.803 | 0.800 | 0.732 | 0.722 |
Rare species: Chao1-N1 | 0 | 3 | 4 | 0 | 4 | 5 | 2 | 12 | 14 |
C. Evenness index | |||||||||
Pielou J=(H’/H’max) | 0.914 | 0.922 | 0.915 | 0.835 | 0.893 | 0.896 | 0.780 | 0.880 | 0.880 |
E. Dominance index | |||||||||
IBP = nmax/n | 0.429 | 0.166 | 0.156 | 0.545 | 0.192 | 0.175 | 0.449 | 0.145 | 0.134 |
Pairwise comparisons of diversity indexes (Student t-test) | |||||||||
Comparison | Shannon-Weaver index H’ | Simpson’s diversity index | |||||||
I vs. II | Nyong: t=-45.93; df=119.29; p=5.7x10-108 * Kienke: t=-76.82; df=961.07; p=0 * | Nyong: t=17.46; df=150.88; p=4.9x10-38 * Kienke: t=31.72; df=709.56; p=3.5x10-138 * | |||||||
Nyong vs. Kienke | Dry season: t=-2.30; df=262.43; p=0.022 * Rainy season: t=-5.28; df=4,285.20; p=1.3x10-7 * Both seasons: t=-3.62; df=4,316.60; p=3.0x10-4 * | Dry season: t=2.47; df=273.73; p=0.014 * Rainy season: t=4.46; df=4,615.20; p=8.3x10-6 * Both seasons: t=2.95; df=4,613.70; p=0.003 * | |||||||
SAD model | AIC (BIC) and the best fitted model | |||||
|---|---|---|---|---|---|---|
Nyong River mouth | Kienke River mouth | |||||
Dry season 145.8x105 cells 3 species | Rainy season 2,312.5x105 cells 17 species | Pooled data 2,457.3x105 cells 20 species | Dry season 687.5x105 cells 3 species | Rainy season 7,281.3x105 cells 17 species | Pooled data 7,968.8x105 cells 20 species | |
Broken-Stick (BS) | 37.34 (37.34) | 299.05 (299.05) | 307.81 (307.81) | 45.25 (45.25) | 245.70 (245.70) | 319.54 (319.54) |
Log-linear (LL) | 34.99 (34.06) | 178.43 (179.26) | 220.08 (221.08) | 58.29 (57.39) | 283.22 (284.06) | 369.49 (370.48) |
Log-normal (LN) | 30.32 (28.51) * | 153.27 (154.94) * | 184.82 (186.81) * | 53.04 (51.24) * | 226.26 (227.92) * | 262.82 (264.81) * |
Zipf (Z) | 34.21 (32.41) | 214.60 (216.27) | 252.86 (254.85) | 77.00 (75.20) | 627.03 (628.70) | 688.24 (690.23) |
Zipf-Mandelbrot (ZM) | 32.32 (29.61) | 153.67 (156.17) | 186.35 (189.34) | 55.04 (52.34) | 266.80 (269.30) | 347.75 (350.74) |
SAD model | AIC (BIC) and the best fitted model | ||
|---|---|---|---|
Pooled rivers | |||
Dry season n=833.3x105 cells S=6 species | Rainy season n=9,593.8x105 cells S=34 species | Pooled data n=10,427.1x105 cells S=40 species | |
Broken-Stick (BS) | 106.49 (106.49) | 529.39 (529.39) | 630.97 (630.97) |
Log-linear (LL) | 93.65 (93.44) | 583.01 (584.53) | 718.18 (719.87) |
Log-normal (LN) | 96.72 (96.30) | 350.85 (353.90) * | 431.95 (435.33) * |
Zipf (Z) | 109.78 (109.37) | 845.13 (848.18) | 1,039.65 (1043.03) |
Zipf-Mandelbrot (ZM) | 92.41 (91.79) * | 579.59 (584.17) | NA |
Dry season | Rainy season | Pooled seasons | |||||||
|---|---|---|---|---|---|---|---|---|---|
High tide | Low tide | Pooled tides | High tide | Low tide | Pooled tides | High tide | Low tide | Pooled tides | |
A. Overall data sets | |||||||||
Dry season | |||||||||
High tide | 1.000 | ||||||||
Low tide | 0.0 | 1.000 | |||||||
Pooled tides | 1.000 | 0.0 | 1.000 | ||||||
Rainy season | |||||||||
High tide | 0.0 | 0.0 | 0.0 | 1.000 | |||||
Low tide | 0.0 | 0.0 | 0.0 | 0.0 | 1.000 | ||||
Pooled tides | 0.0 | 0.0 | 0.0 | 0.313 | 0.898 | 1.000 | |||
Pooled seasons | |||||||||
High tide | 0.431 | 0.0 | 0.431 | 0.841 | 0.0 | 0.295 | 1.000 | ||
Low tide | 0.0 | 0.0 | 0.0 | 0.0 | 1.000 | 0.898 | 0.0 | 1.000 | |
Pooled tides | 0.123 | 0.0 | 0.123 | 0.295 | 0.864 | 0.966 | 0.386 | 0.864 | 1.000 |
B. Nyong River mouth | |||||||||
Dry season | |||||||||
High tide | 1.000 | ||||||||
Low tide | 0.0 | 1.000 | |||||||
Pooled tides | 0.727 | 0.600 | 1.000 | ||||||
Rainy season | |||||||||
High tide | 0.0 | 0.0 | 0.0 | 1.000 | |||||
Low tide | 0.0 | 0.0 | 0.0 | 0.0 | 1.000 | ||||
Pooled tides | 0.0 | 0.0 | 0.0 | 0.600 | 0.727 | 1.000 | |||
Pooled seasons | |||||||||
High tide | 0.145 | 0.0 | 0.137 | 0.959 | 0.0 | 0.585 | 1.000 | ||
Low tide | 0.0 | 0.087 | 0.082 | 0.0 | 0.977 | 0.715 | 0.0 | 1.000 | |
Pooled tides | 0.066 | 0.050 | 0.113 | 0.574 | 0.699 | 0.969 | 0.608 | 0.721 | 1.000 |
C. Kienke River mouth | |||||||||
Dry season | |||||||||
High tide | 1.000 | ||||||||
Low tide | 0.0 | 1.000 | |||||||
Pooled tides | 0.706 | 0.625 | 1.000 | ||||||
Rainy season | |||||||||
High tide | 0.0 | 0.0 | 0.0 | 1.000 | |||||
Low tide | 0.0 | 0.0 | 0.0 | 0.0 | 1.000 | ||||
Pooled tides | 0.0 | 0.0 | 0.0 | 0.151 | 0.957 | 1.000 | |||
Pooled seasons | |||||||||
High tide | 0.558 | 0.0 | 0.453 | 0.760 | 0.0 | 0.144 | 1.000 | ||
Low tide | 0.0 | 0.086 | 0.081 | 0.0 | 0.977 | 0.936 | 0.0 | 1.000 | |
Pooled tides | 0.090 | 0.076 | 0.159 | 0.139 | 0.913 | 0.955 | 0.217 | 0.935 | 1.000 |
Species 1/Species 2 | τ (p-value) | Species 1/Species 2 | τ (p-value) | Species 1/Species 2 | τ (p-value) | |||
|---|---|---|---|---|---|---|---|---|
A. Overall pooled data from both seasons and both river mouths (n=21) | ||||||||
Ac. exiguoides | Cs. rudolfi | De. thermalis (continued) | ||||||
Co. placentula | 0.737(3.0x10-6) * | Fa. construens | 0.447(0.005) * | Ni. tryblionella | 0.425(0.007) * | |||
Am. ovalis | Cr. erosa | Di. sertularia | ||||||
Cm. apiculata | 0.474 (0.003) * | Cc. meneghiniana | 0.445 (0.005) * | Gn. mutica | 0.364 (0.021) * | |||
Cs. rudolfi | 0.322 (0.041) * | De. elegans | 0.445 (0.005) * | Ni. amphibia | 0.568 (3.2x10-4) * | |||
Gn. mutica | 0.742(2.5x10-6) * | Rh. longiseta | 0.360 (0.023) * | Go. olivaceum | ||||
Ca. noricus | Cc. meneghiniana | Rh. longiseta | 0.536 (0.001) * | |||||
Ch. muelleri | 0.520 (0.001) * | Cc. stelligera | 0.497 (0.002) * | Gn. mutica | ||||
Ce. hirundinella | Cm. apiculata | Pi. cardinaliculus | 0.645 (4.4x10-5) * | |||||
Cr. erosa | 0.431 (0.006) * | Di. sertularia | 0.568(3.2x10-4)* | Ni. amphibia | ||||
Cc. meneghiniana | 0.378 (0.016) * | Gn. mutica | 0.716(5.6x10-6)* | Ni. tryblionella | 0.533 (0.001) * | |||
Ch. muelleri | Cm. solea | Pi. cardinaliculus | ||||||
Go. olivaceum | 0.424 (0.007) * | De. elegans | 0.645(4.4x10-5)* | St. astraea | 0.424 (0.007) * | |||
Ni. amphibia | 0.592(1.7x10-4) * | Ni. sigma | 0.716(5.6x10-6)* | |||||
Rh. longiseta | 0.332 (0.035) * | De. elegans | ||||||
Co. placentula | Rh. longiseta | 0.379 (0.016) * | ||||||
Cm. apiculata | 0.598(1.5x10-4) * | De. thermalis | ||||||
Gn. mutica | 0.385 (0.015) * | Gn. gigas | 0.328 (0.037) * | |||||
B. Nyong River mouth (n=12) | ||||||||
Ac. exiguoides | Cs. rudolfi (continued) | Cm. solea | ||||||
Co. placentula | 0.647 (0.006) * | Fa. construens | 0.671 (0.004) * | De. elegans | 0.580 (0.013) * | |||
Am. ovalis | Cr. erosa | Ni. sigma | 0.725 (0.002) * | |||||
Cm. apiculata | 0.580 (0.013) * | De. elegans | 0.725 (0.002) * | De. elegans | ||||
Gn. mutica | 0.895 (1.3x10-4) * | Ni. amphibia | 0.580 (0.013) * | Rh. longiseta | 0.725 (0.002) * | |||
Pi. cardinaliculus | 0.725 (0.002) * | Rh. longiseta | 0.474 (0.043) * | Di. sertularia | ||||
Ca. noricus | Cc. stelligera | Gn. mutica | 0.474 (0.043) * | |||||
Cs. rudolfi | 0.580 (0.013) * | Cm. solea | 0.725 (0.002) * | Fa. construens | ||||
Ce. hirundinella | Cm. apiculata | Ni. amphibia | 0.671 (0.004) * | |||||
De. elegans | 0.671 (0.004) * | Di. sertularia | 0.725 (0.002) * | Gn. mutica | ||||
Co. placentula | Gn. mutica | 0.725 (0.002) * | Pi. cardinaliculus | 0.580 (0.013) * | ||||
Cm. apiculata | 0.620 (0.008) * | Cm. apiculata | ||||||
Cs. rudolfi | Rh. longiseta | 0.580 (0.013) * | ||||||
Di. sertularia | 0.580 (0.013) * | |||||||
C. Kienke River mouth (n=9) | ||||||||
Am. ovalis | Cs. rudolfi | Di. sertularia | ||||||
Cs. rudolfi | 1.000 (1.7x10-4) * | De. thermalis | 0.730 (0.006) * | Ni. amphibia | 0.548 (0.040) * | |||
De. thermalis | 0.730 (0.006) * | Cr. erosa | Ni. tryblionella | 1.000 (1.7x10-4) * | ||||
Ca. noricus | Rh. longiseta | 0.553 (0.038) * | Go. olivaceum | |||||
Ch. muelleri | 1.000 (1.7x10-4) * | Cc. stelligera | Rh. longiseta | 0.789 (0.003) * | ||||
Ni. amphibia | 0.730 (0.006) * | Di. sertularia | 0.730 (0.006) * | Ni. amphibia | ||||
Ce. hirundinella | Ni. tryblionella | 0.730 (0.006) * | Pi. cardinaliculus | 0.548 (0.040) * | ||||
St. astraea | 0.617 (0.021) * | De. thermalis | ||||||
Ch. muelleri | Di. sertularia | 0.548 (0.040) * | ||||||
Ni. amphibia | 0.730 (0.006) * | Ni. tryblionella | 0.548 (0.040) * | |||||
| [1] | Guiry, M. D. How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing. Journal of Phycology. 2024, 00, 1–15. |
| [2] | Cavalier-Smith, T. Eukaryote kingdoms: seven or nine? Biosystems. 1981, 14, 461-481. |
| [3] | Cavalier-Smith, T. Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European Journal of Protistology. 2013, 49, 115-178. |
| [4] | Cavalier-Smith, T. Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma. 2018, 255, 297-357. |
| [5] | Aubert, D. Une nouvelle mégaclassification pragmatique du vivant. Médecine/Sciences. 2016, 32, 497-499. |
| [6] | Johnson, L. R., John, D. M., Whitton, B. A., Brook, A. J. Phylum Xanthophyta (Tribophyta) (Yellow-Green Algae). In The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae, Second Edition, Johnson, L. R., Ed., Cambridge University Press. 2021, pp. 318-345. |
| [7] | Guiry, M. D. How many species of algae are there? Journal of Phycology. 2012, 48(5), 1057–1063. |
| [8] | Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., Cavalier-Smith, T., Guiry, M. D., and, Kirk, P. M. A Higher Level Classification of All Living Organisms. PLoS ONE. 2015, 10(4), e0119248. |
| [9] | Hagen, J. Five Kingdoms, More or Less: Robert Whittaker and the Broad Classification of Organisms. BioScience, 2012, 62. 67-74. |
| [10] | Integrated Taxonomic Information System. “Chromista". Available from www.itis.gov, CC0). [Accessed 29 Febuary 2024], |
| [11] |
DiatomBase. “Chromista”. Available from
https://diatombase.org [Accessed 26 Febuary 2024]. |
| [12] | Bloch, K., Vardi, P. Microalgae as photosynthetic oxygen generators for pollution control, life support systems and medicine, In Algae: Nutrition, Pollution Control and Energy Sources, Hagen, K. N., Ed., Nova Science Publishers, Inc. Tel-Aviv University, Petah Tikva, Israel; 2009, pp. 3-11. |
| [13] | Zalat, A. A., Nitychoruk, J., Chodyka, M., Pidek, I. A., Welc, F. Recent and fossil freshwater diatoms of Poland: taxonomy, distribution and their significance in the environmental reconstruction. Part 1. Coscinodiscophyceae, Mediophyceae and Fragilariophycidae. John Paul II University of Applied Sciences, Biała Podlaska; 2022, pp. 1-306 |
| [14] | Jahn, R., Kusber, W. H., Romero, O. E. Cocconeis pediculus Ehrenberg and C. placentula Ehrenberg var. placentula (Bacillariophyta): Typification and taxonomy. Fottea. 2009, 9(2), 275–288. |
| [15] | Hecky, R. E., Kilhan, P. Diatoms in Alkaline, Saline Lakes: Ecology and Geochemical Implications. Limnology and Oceanography. 1973, 18(1), 53-71. |
| [16] | Spaulding, S. A., Potapova, M. G., Bishop, I. W., Lee, S. S., Gasperak, T. S., Jovanovska, E., Edlund, M. B.. Diatoms.org: supporting taxonomists, connecting communities. Diatom Research. 2021, 36(4): 291-304. |
| [17] | Houk, V.; Klee, R., Tanaka, H. Atlas of freshwater centric diatoms with a brief key and descriptions, Part III, Stephanodiscaceae A, Cyclotella, Tertiarius, Discostella. Fottea. 2010, 10(Supplement), 1–498. |
| [18] | Krammer, K., Lange-Bertalot, H. Bacillariophyceae, 2nd part: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa, Band 2/2. Ettl, H., Gerloff, J., Heynıg, H., Mollenhauer, D., Eds., Gustav Fischer Verlag, Stuttgart, Germany; 1988, pp. 1-596. |
| [19] | Krammer, K, Lange-Bertalot, H. Bacillariophyceae, 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süsswasserflora Von Mitteleuropa, Band 2/3. Ettl, H., Gerloff, J., Heynıg, H., Mollenhauer, D., Eds., Gustav Fischer Verlag, Stuttgart, Germany; 1991, pp. 1-596. |
| [20] | Mohanty, T. R.,•Tiwari, N. K.,•Kumari, S.,•Ray, A.,•Manna, R. K., Bayen, S.,•Roy, S.,•Gupta, S. D.,•Ramteke, M. H.,•Swain, H. S., Bhor, M.,•Das, B. K. Variation of Aulacoseira granulata as an eco pollution indicator in subtropical large river Ganga in India: a multivariate analytical approach. Environmental science and pollution research international. 2022, 29(25), 37498-37512 |
| [21] | Taylor, J. C., Harding, W. R., Archibald, C. G. M. An Illustrated Guide to Some Common Diatom Species from South Africa. WRC Report TT 282/07. Water Research Commission. Gezina, Pretoria, South Africa; 2007, pp. 1-225. |
| [22] |
DeColibus, D. “Tabellaria flocculosa (Roth) Kütz. 1844. In Diatoms of North America”. Available from
https://diatoms.org [Accessed 01 March 2024]. |
| [23] |
Lowe, R. “Epithemia turgida. In Diatoms of North America”. Available from
https://diatoms.org/species/epithemia_turgida [Accessed 22 January 2024]. |
| [24] | Mojiri, A, Baharlooeian, M, Zahed M. A. The Potential of Chaetoceros muelleri in Bioremediation of Antibiotics: Performance and Optimization. International Journal of Environmental Research in Public Health. 2021, 22, 18(3), 977. |
| [25] | Sivonen, K. Cyanobacterial toxins and toxin production. Phycologia. 1996, 35, 12-24. |
| [26] | Mankiewicz, J., Tarczynska, M., Walter, Z., Zalewski, M. Natural Toxins from Cyanobacteria, Acta Biologica Cracoviensia Series Botanica. 2003, 45(2), 9-20. |
| [27] | WHO (World Health Organization). Chapter 8 – Algae and Cyanobacteria in Freshwater. In Guidelines for safe recreational waters. Volume 1 – Coastal and fresh waters. Geneva, Switzerland; 2003, pp. 136-158. |
| [28] | Zanchett, G., Oliveira-Filho, E. Cyanobacteria and Cyanotoxins: From Impacts on Aquatic Ecosystems and Human Health to Anticarcinogenic Effects. Toxins. 2013, 5, 1896-917. |
| [29] | Rastogi, R. P., Madamwar, D., Incharoensakdi, A. Bloom Dynamics of Cyanobacteria and Their Toxins: Environmental Health Impacts and Mitigation Strategies. Frontiers in Microbiology. 2015, 6, 1254. |
| [30] | Beasley, V. R. Harmful Algal Blooms (Phycotoxins). In Reference Module in Earth Systems and Environmental Sciences, Elsevier Inc. 2019. |
| [31] | Nicholls, K. H., Kennedy, W., Hammett, C. A fish-kill in Heart Lake, Ontario, associated with the collapse of a massive population of Ceratium hirundinella (Dinophyceae). Freshwater Biology. 1980, 10, 553-561. |
| [32] | Caturao, R. D. Harmful and toxic algae. In Health Management in Aquaculture (2nd edition). Aquaculture Department, Southeast Asian Fisheries Development Center, Lio-Po G. D. & Inui, Y., Eds., Tigbauan, Iloilo, Philippines; 2010, pp. 170-182. |
| [33] | Magray, A. R., Lone, S. A., Ganai, B. A., Ahmad, F., Hafeez, S. The first detection and in vivo pathogenicity characterization of Saprolegnia delica from Kashmir Himalayas. Aquaculture. 2021, 542, 736876. |
| [34] | Raghukumar, S. Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids). European Journal of Protistology. 2022, 38(2), 127-145. |
| [35] | Brummett, R. E., Nguenga, D., Tiotsop, F., Abina, J.-C... The commercial fishery of the middle Nyong River, Cameroon: productivity and environmental threats. Smithiana Bulletin. 2010, 11, 3-16. |
| [36] | Olivry, J. C. Monographie hydrologique. Fleuves et rivières du Cameroun. Ministère de l’Enseignement Supérieur et de la Recherche Scientifique au Cameroun - Institut Français de Recherche Scientifique pour le Developpement en Coopération/ (MESRES-ORSTOM). Collection "Monographies Hydrologiques ORSTOM" No 9, Paris. 1986, pp. 1-781. |
| [37] | Dounias, E., Cogels, S., Mvé Mbida, S., Carrière, S. The safety net role of inland fishing in the subsistence strategy of multiactive forest dwellers in southern Cameroon. Revue d’Ethnoécologie. 2016, 10, 11-56. |
| [38] | Mama, A. C., Mbeng, O., Dongmo, C., Ohondja, A. Tidal Variations and its impacts on the abundance and diversity of phytoplankton in the Nyong estuary of Cameroon. Journal of Multidisciplinary Engineering Science and Technology. 2016, 3(1), 3159-3199. |
| [39] | Mama, A. C., Youbouni Ghepdeu, G. F., Ngoupayou Ndam, J. R., Bonga, M. D., Onana, F. M., Onguene, R. Assessment of water quality in the lower Nyong estuary (Cameroon, Atlantic Coast) from environmental variables and phytoplankton communities’ composition. African Journal of Environmental Science and Technology. 2018, 12(6), 198-208. |
| [40] | Essomba Biloa, R. E., Noah Ewoti, O. V., Tuekam Kayo, R. P., Sob Nangou, P. B., Tchakounté, S., Onana, F. M., Nyamsi Tchatcho, N. L., Zebaze Togouet S. H. Zooplankton Dynamics of the Kienke Estuary (Kribi, South Region of Cameroon): Importance of Physico-Chemical Parameters. Open Journal of Ecology. 2021, 11, 837-869. |
| [41] | Mokam, C. C., Kenne Toukem, A. S., Dongmo Teufack, C., Amougou Dzou, F. T., Tsekane, S. J., Moukhtar, M., Mbianda, A. P., Kenne M. Water Quality, Biodiversity and Abundance of Blue-Green Algae in Nyong and Kienke River Mouths (South-Cameroon). International Journal of Ecotoxicology and Ecobiology. 2024, 9(1), 1-27. |
| [42] |
Climate-Data.org. Climat Kribi (Cameroun). Available from
https://fr.climate-data.org [Accessed 22 January 2024]. |
| [43] | Compère, P. Algues de la région du Lac Tchad. IV: Diatomophycées. Cahier ORSTOM, série. Hydrohiologie. 1975, 9(4), 205-290. |
| [44] | Carty, S. Freshwater Dinoflagellates of North America. Comstock Publishing Associates, Cornell University Press. Ithaca, New York, USA. 2014, pp. 280. |
| [45] | Olenina, I., Hajdu, S., Edler, L., Andersson, A., Wasmund, N., Busch, S., Göbel, J., Gromisz, S., Huseby, S., Huttunen, M., Jaanus, A., Kokkonen, P., Ledaine, I., Niemkiewicz, E.. Biovolumes and size-classes of phytoplankton in the Baltic Sea HELCOM Balt. Sea Environ. Proc. No. 106. 2006, pp. 1-144. |
| [46] | Bellinger, E. G., Sigee, D. C. Freshwater Algae. Identification and Use as Bioindicators. Wiley-Blackwell, Hoboken, USA. 2010, pp. 1-271. |
| [47] | Jahn, R., Kusber, W.-H. A Key to Diatom Nomenclature. Diatom Research. 2009, 24(1), 101-111. |
| [48] |
DREAL (Direction Régionale de l’Environnement, de l’Aménagement et du logement d’Occitanie). "Atlas des Diatomées de l’ex partie Languedoc-Roussillon. Languedoc Roussillon". Available from
https://www.occitanie.developpement-durable.gouv.fr/atlas-des-diatomees-de-l-ex-partie-languedoc-a25336.html [Accessed 22 January 2021]. |
| [49] | Sonneman, J. A., Sincock, A., Fluin, J., Reid, M., Newall, P., Tibby, J., Gell, P. An Illustrated Guide to Common Stream Diatom Species from Temperate Australia. Cooperative Research Centre for Freshwater Ecology. Identification guide n°33. Land & Water Resources Research & Development Corporation. 2nd Australian Algal Workshop, Adelaide University, 17-19th April, 2000, pp. 1-166. |
| [50] | Kumar, D. S., Maurya, O. N. Floristic Survey of Algae in Vikramsila Gangetic Dolphin Sanctuary, Bihar (India). Nelumbo. 2015, 57, 124-134. |
| [51] |
WoRMS (World Register of Marine Species). “Chromista”. Available from
https://www.marinespecies.org/aphia.php?p=taxdetails&id=7 at VLIZ. [Accessed 06 February 2024]. |
| [52] |
Guiry, M. D., Guiry, G. M. 2024. “Kingdom Chromista Cavalier-Smith 1981”. AlgaeBase. World-wide electronic publication, University of Galway, Ireland. Available from
https://www.algaebase.org [Accessed 28 February 2024]. |
| [53] | Wilson, J. B. Methods for fitting dominance/diversity curves. Journal of Vegetation Science. 1991, 2(1), 35-46. |
| [54] | Iganaki, H. Mise au point de la loi de Motomura et essai d’une ecologie ́evolutive. Vie et Milieu. 1967, 18, 153-166. |
| [55] | Li, W. Zipf's Law Everywhere. Glottometrics. 2002, 5, 14-21. |
| [56] | Zipf, G. K.,. Human Behavour and the Principle of Least Effort: An introduction to human ecology. 2nd edition, Hafner, New York, NY, USA. 1965, pp. 1-573. |
| [57] | Le, D.-H., Pham, C.-K., Nguyen, T. T. T., Bui, T. T. Parameter extraction and optimization using Levenberg-Marquardt algorithm,. In Proceedings of 2012 IEEE conference. Fourth International Conference on Communications and Electronics (ICCE), Hanoi University of Science and Technology, Hanoi (Vietnam), 2022; pp. 434-437. |
| [58] | Frontier, S. Applications of Fractal Theory to Ecology. In Developments in Numerical Ecology., Legendre P., Legendre, L., Eds., NATO ASI Series book series (volume 14). Springer, Berlin, Heidelberg. 1987, pp. 335-378. |
| [59] | Bach, P., Amanieu, M., Lam-Hoai, T., Lasserre, G. Application du modele de distribution d'abondance de Mandelbrot a l'estimation des captures dans l'étang de Thau. Journal du Conseil/Conseil Permanent International pour l'Exploration de la Mer. 1988, 44, 235-246. |
| [60] | Murthy, Z. V. P. Nonlinear Regression: Levenberg-Marquardt Method, In Encyclopedia of Membranes, Drioli, E., Giorno L., Eds., Springer-Verlag, Berlin, Heidelberg, 2014, pp. 1-3. |
| [61] | Ludwig, J. A., Raynolds, J. F. Statistical Ecology. John Wiley & Sons, New York, USA. 1988, pp. 1-337. |
| [62] |
Friedman, N. & Potapova, M. “Pinnularia cardinaliculus”. In Diatoms of North America. Available from
https://diatoms.org [Accessed 22 January 2024]. |
| [63] | Lange-Bertalot, H., Fuhrmann, A. Contribution to the genus Diploneis (Bacillariophyta): Twelve species from Holarctic freshwater habitats proposed as new to science. Fottea. 2016, 16(2), 157–183. |
| [64] | Kelly, M. G., Whitton, B. A.. The trophic diatom index: a new index for monitoring eutrophication in rivers. Journal of Applied Phycology. 1995, 7, 433-444. |
| [65] | Tokatli, C. Evaulation of water quality by using trophic diatom index: example of Porsuk Dam Lake. Journal of Applied Biological Sciences 2013, 7(1), 01-04. |
| [66] | Kelly, M. G., Adams, C., Graves, A. C., Jamieson, J., Krokowski, J., Lycett, E. B., Murray-Bligh, J., Pritchard, S., Wilkins, C. The trophic diatom index: A User’s manual. Revised Edition. R & D technical Report E2/TR2. Environment Agency, Almondsbury, Bristol, BS32 4UD; 2001, pp. 1-135. |
| [67] | Solak, C., N. The Application of Diatom Indices in the Upper Porsuk Creek Kütahya – Turkey. Turkish Journal of Fisheries and Aquatic Sciences. 2011, 11, 31-36. |
| [68] | Ndjouondo, G. P., Nwamo, R. D., Choula Tegantchouang, F. Diversity and Structure of Microalgae in the Mezam River (Bamenda, Cameroon). African Journal of Environment and Natural Science Research. 2023, 6(1), 19-35. |
| [69] | Menye, D. É., Zébazé Togouet, S. H., Menbohan, S. F., Kemka, N., Nola, M., Boutin, C., Nguetsop, V. F., Djaouda, M., Njiné, T. Bio-écologie des diatomées épilithiques de la rivière Mfoundi (Yaoundé, Cameroun): diversité, distribution spatiale et influence des pollutions organiques. Revue des sciences de l’eau/Journal of Water Science. 2012, 25(3), 203–218. |
| [70] | Castel, J., Courties, C. Composition and differential distribution of zooplankton in Arcachon Bay. Journal of Plankton Research. 1982, 4(3), 417-433. |
| [71] | Kenne, E. L., Yetchom-Fondjo, J. A., Moumite, M. B., Tsekane, S. J., Ngamaleu-Siewe, B., Fouelifack-Nintidem, B., Biawa-Kagmegni, M., Tuekam Kowa, P. S., Fantio, R. M., Yomon, A. K., Kentsop-Tsafong, R. M., Dim-Mbianda, A. M., Kenne M. Species composition and diversity of freshwater snails and land snails at the swampy areas and streams edges in the urban zone of Douala, Cameroon. International Journal of Biological and Chemical Sciences. 2021, 15(4), 1297-1324. |
| [72] | Shea, K., Chesson, P. Community ecology theory as a framework for biological invasions. Trends in Ecology and Evolution. 2002, 17(4), 170-176. |
| [73] | Ferreira, F. C., Petrere-Jr., M. Comments about some species abundance patterns: classic, neutral, and niche partitioning models, Brazilian Journal of Biology, 2008, 68(4, Suppl.), 1003-1012. |
APA Style
Mokam, C. C., Toukem, A. S. K., Teufack, C. D., Dzou, F. T. A., Tsekane, S. J., et al. (2024). Biodiversity and the Community Structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River Mouths (South-Cameroon). International Journal of Ecotoxicology and Ecobiology, 9(1), 28-55. https://doi.org/10.11648/j.ijee.20240901.12
ACS Style
Mokam, C. C.; Toukem, A. S. K.; Teufack, C. D.; Dzou, F. T. A.; Tsekane, S. J., et al. Biodiversity and the Community Structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River Mouths (South-Cameroon). Int. J. Ecotoxicol. Ecobiol. 2024, 9(1), 28-55. doi: 10.11648/j.ijee.20240901.12
AMA Style
Mokam CC, Toukem ASK, Teufack CD, Dzou FTA, Tsekane SJ, et al. Biodiversity and the Community Structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River Mouths (South-Cameroon). Int J Ecotoxicol Ecobiol. 2024;9(1):28-55. doi: 10.11648/j.ijee.20240901.12
@article{10.11648/j.ijee.20240901.12,
author = {Christelle Chimene Mokam and Andrea Sarah Kenne Toukem and Christian Dongmo Teufack and Fabien Tresor Amougou Dzou and Sedrick Junior Tsekane and Mohammadou Moukhtar and Auguste Pharaon Mbianda and Martin Kenne},
title = {Biodiversity and the Community Structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River Mouths (South-Cameroon)
},
journal = {International Journal of Ecotoxicology and Ecobiology},
volume = {9},
number = {1},
pages = {28-55},
doi = {10.11648/j.ijee.20240901.12},
url = {https://doi.org/10.11648/j.ijee.20240901.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijee.20240901.12},
abstract = {A survey was undertaken from March to June 2014 on the biodiversity and the community structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River mouths (South-Cameroon). In each river, raw waters were collected from upstream to downstream at four sites. Cells were counted using the Malassez cells procedure and species were identified. A total of 10427.1x105 cells corresponded to three phyla, eight classes, 23 orders, 32 genera and 40 species (24 freshwater species (60.0% of total species richness and total collection respectively), three marine species (7.5% and 2.4% of the total species richness; and total collection respectively), and one brackish water specialist in Kienke (2.5% and 5.1%), 13 tolerant species (32.5% and 32.6%)). The trophic diatom index revealed undisturbed conditions with no or little alteration of human origin and a low organic pollution (oligotrophic or mesotrophic state) (Nyong: TDI=52.7; Kienke: TDI=69.7; pooled assemblage: TDI=65.0). A low species richness was detected (richness ratio in Nyong: d=0.008; Kienke: d=0.003; pooled rivers: d=0.004), a high species diversity (Shannon index close to maximum) (Nyong: H’=2.742 and H’max=2.996; Kienke: H’=2.685 and H’max=2.996; pooled rivers: H’=3.245 and H’max=3.689), a very low dominance by a few species (Berger-Parker index close to 0) (Nyong: IBP=0.156; Kienke: IBP=0.175; pooled rivers: IBP=0.134), and Hill’s ratio were close to 1 (Nyong: Hill=0.819; Kienke: Hill=0.803; pooled rivers: Hill=0.722). The community was highly even with a high value of the Pielou’s evenness close to 1 (Nyong: J=0.915; Kienke: J=0.896; pooled rivers: J=0.880). Two useful species and one harmful species to fish were rare in Kienke. Species exhibited in Kienke and pooled data in rainy season, a positive global net association while it was negative in Nyong. Assemblage fitted Preston’s model in Nyong with a high environmental constant in the dry season (m’=1.469), low constant in the rainy season (m’=0.947) and the pooled seasons (m’=0.853). In Kienke constants were low (dry season: m’=0.574; rainy season: m’=0.566; pooled seasons: m’=0.581) suggesting a evolved community in less disturbed environments where the majority of species showed moderate abundances. In the dry season, the pooled assemblage functionned on the basis of maintaining a complex information network (close to ecological balance) developed at spatio-temporal scales (ZM model) and it presented a low force of regeneration (fractal dimension of the distribution of individuals among species (1/γ)=0.925<1). The evolved oligotrophic state (close to natural balance) of the chromists’ community should be preserved and protected and the studied rivers classified as reference.
},
year = {2024}
}
TY - JOUR T1 - Biodiversity and the Community Structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River Mouths (South-Cameroon) AU - Christelle Chimene Mokam AU - Andrea Sarah Kenne Toukem AU - Christian Dongmo Teufack AU - Fabien Tresor Amougou Dzou AU - Sedrick Junior Tsekane AU - Mohammadou Moukhtar AU - Auguste Pharaon Mbianda AU - Martin Kenne Y1 - 2024/04/02 PY - 2024 N1 - https://doi.org/10.11648/j.ijee.20240901.12 DO - 10.11648/j.ijee.20240901.12 T2 - International Journal of Ecotoxicology and Ecobiology JF - International Journal of Ecotoxicology and Ecobiology JO - International Journal of Ecotoxicology and Ecobiology SP - 28 EP - 55 PB - Science Publishing Group SN - 2575-1735 UR - https://doi.org/10.11648/j.ijee.20240901.12 AB - A survey was undertaken from March to June 2014 on the biodiversity and the community structure of Chromista Cavalier-Smith, 1981 in Nyong and Kienke River mouths (South-Cameroon). In each river, raw waters were collected from upstream to downstream at four sites. Cells were counted using the Malassez cells procedure and species were identified. A total of 10427.1x105 cells corresponded to three phyla, eight classes, 23 orders, 32 genera and 40 species (24 freshwater species (60.0% of total species richness and total collection respectively), three marine species (7.5% and 2.4% of the total species richness; and total collection respectively), and one brackish water specialist in Kienke (2.5% and 5.1%), 13 tolerant species (32.5% and 32.6%)). The trophic diatom index revealed undisturbed conditions with no or little alteration of human origin and a low organic pollution (oligotrophic or mesotrophic state) (Nyong: TDI=52.7; Kienke: TDI=69.7; pooled assemblage: TDI=65.0). A low species richness was detected (richness ratio in Nyong: d=0.008; Kienke: d=0.003; pooled rivers: d=0.004), a high species diversity (Shannon index close to maximum) (Nyong: H’=2.742 and H’max=2.996; Kienke: H’=2.685 and H’max=2.996; pooled rivers: H’=3.245 and H’max=3.689), a very low dominance by a few species (Berger-Parker index close to 0) (Nyong: IBP=0.156; Kienke: IBP=0.175; pooled rivers: IBP=0.134), and Hill’s ratio were close to 1 (Nyong: Hill=0.819; Kienke: Hill=0.803; pooled rivers: Hill=0.722). The community was highly even with a high value of the Pielou’s evenness close to 1 (Nyong: J=0.915; Kienke: J=0.896; pooled rivers: J=0.880). Two useful species and one harmful species to fish were rare in Kienke. Species exhibited in Kienke and pooled data in rainy season, a positive global net association while it was negative in Nyong. Assemblage fitted Preston’s model in Nyong with a high environmental constant in the dry season (m’=1.469), low constant in the rainy season (m’=0.947) and the pooled seasons (m’=0.853). In Kienke constants were low (dry season: m’=0.574; rainy season: m’=0.566; pooled seasons: m’=0.581) suggesting a evolved community in less disturbed environments where the majority of species showed moderate abundances. In the dry season, the pooled assemblage functionned on the basis of maintaining a complex information network (close to ecological balance) developed at spatio-temporal scales (ZM model) and it presented a low force of regeneration (fractal dimension of the distribution of individuals among species (1/γ)=0.925<1). The evolved oligotrophic state (close to natural balance) of the chromists’ community should be preserved and protected and the studied rivers classified as reference. VL - 9 IS - 1 ER -
Department of Biology and Physiology of Animal Organisms, Faculty of Science, University of Douala, Douala, Cameroon; Department of Biology, Ecology and Evolution, Liège (Sart-Tilman), Belgium
Department of Biology and Physiology of Animal Organisms, Faculty of Science, University of Douala, Douala, Cameroon
Laboratory of the Specialized Center for Research on Marine Ecosystems, Kribi, Cameroon
Laboratory of the Specialized Center for Research on Marine Ecosystems, Kribi, Cameroon
Department of Biology and Physiology of Animal Organisms, Faculty of Science, University of Douala, Douala, Cameroon
Department of Biology and Physiology of Animal Organisms, Faculty of Science, University of Douala, Douala, Cameroon
Department of Biology of Vegetal Organisms, Faculty of Science, University of Douala, Douala, Cameroon
Department of Biology and Physiology of Animal Organisms, Faculty of Science, University of Douala, Douala, Cameroon
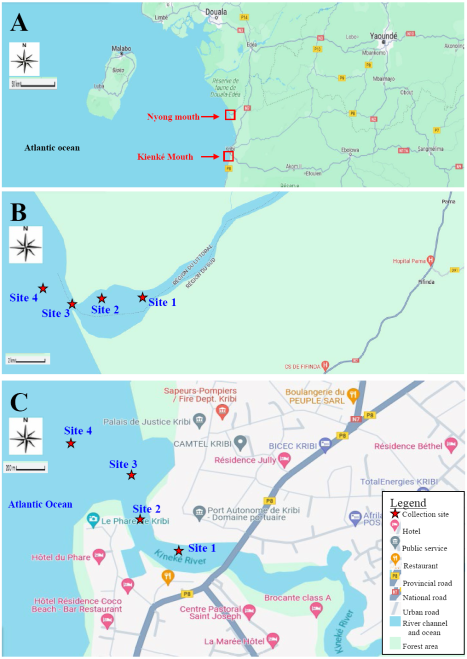
Figure 1. Location of the study sites in Southern coastal zone of Cameroon (southern province, Ocean department). A: Location of the Nyong and Kienke River mouths; B: Location of the collection sites in the Nyong River mouth; C: Location of the collection sites in the Kienke River mouth.
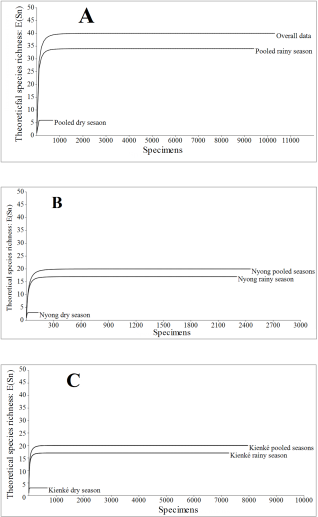
Figure 2. Individual rarefaction curves of the Chromista collections during the dry season and the rainy season in Nyong and Kienke River mouths at both tides.
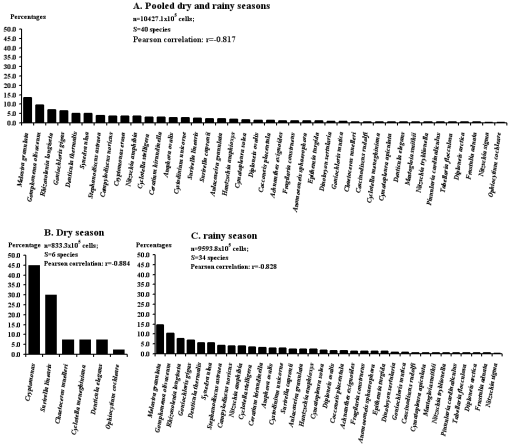
Figure 3. Rank-frequency diagram of the pooled Chromista collection in both river mouths at both tides, showing species in decreasing order of numerical occurrence.
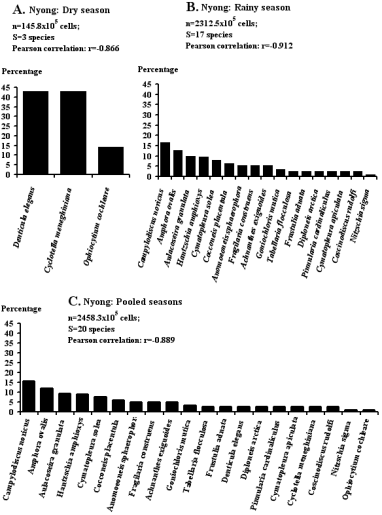
Figure 4. Rank-frequency diagrams of the Chromista collection in the Nyong River mouth at both tides and both seasons, showing species in decreasing order of numerical occurrence.
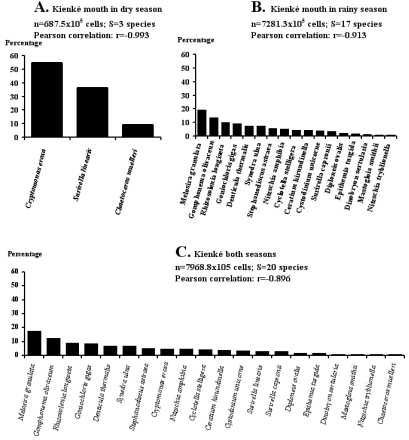
Figure 5. Rank-frequency diagrams of the Chromista collection in the Kienke River mouth at both tides and both seasons, showing species in decreasing order of numerical occurrence.

